
Välise rakumembraani struktuur. Bioloogiliste membraanide struktuur ja funktsioonid

piiritlev ( barjäär) - eraldada raku sisu väliskeskkonnast;
Reguleerida raku ja keskkonna vahelist vahetust;
Jagage rakud sektsioonideks või sektsioonideks, mis on ette nähtud teatud spetsiifiliste metaboolsete radade jaoks ( jagamine);
See on teatud keemiliste reaktsioonide koht (fotosünteesi kerged reaktsioonid kloroplastides, oksüdatiivne fosforüülimine hingamise ajal mitokondrites);
Pakkuda sidet mitmerakuliste organismide kudedes olevate rakkude vahel;
Transport- teostab transmembraanset transporti.
Retseptor- on väliseid stiimuleid ära tundvate retseptorite lokaliseerimise koht.
Ainete transport läbi membraani on membraani üks juhtivaid funktsioone, mis tagab ainete vahetuse raku ja väliskeskkonna vahel. Sõltuvalt ainete ülekande energiakuludest on:
passiivne transport või hõlbustatud difusioon;
aktiivne (selektiivne) transport ATP ja ensüümide osalusel.
transport membraanpakendis. On olemas endotsütoos (rakku) ja eksotsütoos (rakust välja) – mehhanismid, mis transpordivad suuri osakesi ja makromolekule läbi membraani. Endotsütoosi ajal moodustab plasmamembraan invaginatsiooni, selle servad ühinevad ja tsütoplasmasse kinnitub vesiikul. Vesiikul on tsütoplasmast piiratud ühe membraaniga, mis on osa välimisest tsütoplasmaatilisest membraanist. Eristage fagotsütoosi ja pinotsütoosi. Fagotsütoos on suurte, pigem tahkete osakeste imendumine. Näiteks lümfotsüütide, algloomade jne fagotsütoos. Pinotsütoos on vedelikupiiskade hõivamine ja absorbeerimine selles lahustunud ainetega.
Eksotsütoos on erinevate ainete eemaldamise protsess rakust. Eksotsütoosi ajal ühineb vesiikuli või vakuooli membraan välise tsütoplasmaatilise membraaniga. Vesiikuli sisu eemaldatakse rakupinnalt ja membraan lisatakse välimisse tsütoplasmamembraani.
Keskmiselt passiivne laenguta molekulide transport on vesiniku ja laengute kontsentratsioonide vahe, s.o. elektrokeemiline gradient. Ained liiguvad kõrgema gradiendiga alalt madalama gradiendiga alale. Transpordikiirus sõltub gradiendi erinevusest.
Lihtdifusioon on ainete transport otse läbi lipiidide kaksikkihi. Gaasidele iseloomulikud, mittepolaarsed või väikesed laenguta polaarsed molekulid, lahustuvad rasvades. Vesi tungib kiiresti läbi kaksikkihi, sest. selle molekul on väike ja elektriliselt neutraalne. Vee difusiooni läbi membraanide nimetatakse osmoosiks.
Difusioon läbi membraanikanalite on laetud molekulide ja ioonide (Na, K, Ca, Cl) transport, mis tungivad läbi membraani, kuna selles on spetsiaalsed kanaleid moodustavad valgud, mis moodustavad veepoore.
Hõlbustatud difusioon on ainete transport spetsiaalsete transportvalkude abil. Iga valk vastutab rangelt määratletud molekuli või seotud molekulide rühma eest, suhtleb sellega ja liigub läbi membraani. Näiteks suhkrud, aminohapped, nukleotiidid ja muud polaarsed molekulid.
aktiivne transport teostavad valgud - kandjad (ATPaas) elektrokeemilise gradiendi vastu energiakuluga. Selle allikaks on ATP molekulid. Näiteks naatrium-kaaliumpump.
Kaaliumi kontsentratsioon rakus on palju suurem kui väljaspool seda ja naatriumi kontsentratsioon vastupidi. Seetõttu difundeeruvad kaalium- ja naatriumkatioonid passiivselt mööda kontsentratsioonigradienti läbi membraani veepooride. See on tingitud asjaolust, et membraani läbilaskvus kaaliumiioonide jaoks on suurem kui naatriumioonide puhul. Sellest lähtuvalt difundeerub kaalium rakust kiiremini kui naatrium rakku. Raku normaalseks toimimiseks on aga vajalik teatud suhe 3 kaaliumi- ja 2 naatriumiooni. Seetõttu on membraanis naatrium-kaaliumpump, mis pumpab aktiivselt naatriumi rakust välja ja kaaliumi rakku. See pump on transmembraanne membraanivalk, mis on võimeline konformatsioonilisi ümberkorraldusi tegema. Seetõttu võib see enda külge kinnituda nii kaaliumiioone kui ka naatriumiioone (antiport). Protsess on energiamahukas:
Naatriumioonid ja ATP molekul sisenevad pumbavalgusse membraani seest ning kaaliumiioonid väljastpoolt.
Naatriumioonid ühinevad valgu molekuliga ja valk omandab ATPaasi aktiivsuse, st. võime põhjustada ATP hüdrolüüsi, millega kaasneb pumpa käitava energia vabanemine.
ATP hüdrolüüsi käigus vabanev fosfaat kinnitub valgu külge, s.o. fosforüülib valku.
Fosforüülimine põhjustab valgu konformatsioonilise muutuse, see ei suuda säilitada naatriumioone. Nad vabastatakse ja lähevad rakust välja.
Valgu uus konformatsioon soodustab sellele kaaliumioonide lisandumist.
Kaaliumiioonide lisamine põhjustab valgu defosforüülimist. Ta muudab jälle oma kehaehitust.
Valgu konformatsiooni muutus viib kaaliumiioonide vabanemiseni rakus.
Valk on taas valmis naatriumioone enda külge kinnitama.
Ühe töötsükli jooksul pumpab pump rakust välja 3 naatriumiooni ja 2 kaaliumiiooni.
Tsütoplasma- raku kohustuslik komponent, mis on suletud raku pinnaaparaadi ja tuuma vahele. See on kompleksne heterogeenne struktuurikompleks, mis koosneb:
hüaloplasma
organellid (tsütoplasma püsivad komponendid)
kandmised - tsütoplasma ajutised komponendid.
tsütoplasmaatiline maatriks(hüaloplasma) on raku sisemine sisu – värvitu, paks ja läbipaistev kolloidne lahus. Tsütoplasmaatilise maatriksi komponendid viivad rakus läbi biosünteesi protsesse, sisaldavad peamiselt anaeroobse glükolüüsi tõttu energia tekkeks vajalikke ensüüme.
Tsütoplasmaatilise maatriksi põhiomadused.
Määrab raku kolloidsed omadused. Koos vakuolaarsüsteemi rakusiseste membraanidega võib seda pidada väga heterogeenseks või mitmefaasiliseks kolloidsüsteemiks.
Tagab tsütoplasma viskoossuse muutuse, ülemineku geelilt (paksemalt) soolile (vedelam), mis toimub väliste ja sisemiste tegurite mõjul.
Tagab tsüklosi, amööbide liikumise, rakkude jagunemise ja pigmendi liikumise kromatofoorides.
Määrab intratsellulaarsete komponentide asukoha polaarsuse.
Tagab rakkude mehaanilised omadused – elastsuse, ühinemisvõime, jäikuse.
Organellid- püsivad rakustruktuurid, mis tagavad raku poolt spetsiifiliste funktsioonide täitmise. Sõltuvalt struktuuri omadustest on olemas:
membraansed organellid - omavad membraanistruktuuri. Need võivad olla ühemembraanilised (ER, Golgi aparaat, lüsosoomid, taimerakkude vakuoolid). Topeltmembraan (mitokondrid, plastiidid, tuum).
Mittemembraansed organellid - ei oma membraanistruktuuri (kromosoomid, ribosoomid, rakukeskus, tsütoskelett).
Üldotstarbelised organellid – omased kõikidele rakkudele: tuum, mitokondrid, rakukeskus, Golgi aparaat, ribosoomid, ER, lüsosoomid. Kui teatud tüüpi rakkudele on iseloomulikud organellid, nimetatakse neid spetsiaalseteks organellideks (näiteks müofibrillid, mis tõmbavad lihaskiudu kokku).
Endoplasmaatiline retikulum- ühtne pidev struktuur, mille membraan moodustab palju invaginatsioone ja volte, mis näevad välja nagu tuubulid, mikrovakuoolid ja suured tsisternid. EPS membraanid on ühelt poolt seotud raku tsütoplasmaatilise membraaniga ja teiselt poolt tuumamembraani väliskestaga.
EPS-i on kahte tüüpi – kare ja sile.
Karedas või granuleeritud ER-s on tsisternid ja torukesed seotud ribosoomidega. on membraani välimine külg Siledas või agranulaarses EPS-is puudub seos ribosoomidega. See on membraani sisemus.
Kellelegi pole saladus, et kõik meie planeedi elusolendid koosnevad nende rakkudest, nendest lugematutest "" orgaanilistest ainetest. Rakke omakorda ümbritseb spetsiaalne kaitsekesta – membraan, mis mängib raku elus väga olulist rolli ning rakumembraani funktsioonid ei piirdu ainult raku kaitsmisega, vaid kujutavad endast kõige keerulisemat sellega seotud mehhanismi. rakkude paljunemisel, toitumisel ja taastumisel.
Mis on rakumembraan
Sõna "membraan" ise on ladina keelest tõlgitud kui "kile", kuigi membraan ei ole lihtsalt kile, millesse rakk on mähitud, vaid kahe omavahel ühendatud ja erinevate omadustega kile kombinatsioon. Tegelikult on rakumembraan kolmekihiline lipoproteiini (rasvvalk) kest, mis eraldab iga raku naaberrakkudest ja keskkonnast ning teostab kontrollitud vahetust rakkude ja keskkonna vahel, see on raku akadeemiline määratlus. membraan on.
Membraani väärtus on lihtsalt tohutu, sest see mitte ainult ei eralda üht rakku teisest, vaid tagab ka raku koostoime nii teiste rakkudega kui ka keskkonnaga.
Rakumembraanide uurimise ajalugu
Olulise panuse rakumembraani uurimisse andsid kaks Saksa teadlast Gorter ja Grendel juba 1925. aastal. Siis õnnestus neil punaste vereliblede - erütrotsüütidega läbi viia kompleksne bioloogiline eksperiment, mille käigus said teadlased nn "varjud", tühjad erütrotsüütide kestad, mis volditi üheks hunnikuks ja mõõdeti pindala, ja ka arvutas neis sisalduvate lipiidide koguse. Saadud lipiidide hulga põhjal jõudsid teadlased järeldusele, et neist piisab täpselt rakumembraani topeltkihi jaoks.
1935. aastal tegi teine paar rakumembraani-uurijaid, seekord ameeriklased Daniel ja Dawson, pärast mitmeid pikki katseid kindlaks valgusisalduse rakumembraanis. Muidu oli võimatu seletada, miks membraanil on nii suur pindpinevus. Teadlased esitasid nutikalt võileiva kujul oleva rakumembraani mudeli, milles leiva rolli täidavad homogeensed lipiid-valgukihid ja nende vahel või asemel tühjus.
1950. aastal, Danieli ja Dawsoni elektroonilise teooria tulekuga, oli võimalik juba praktilisi tähelepanekuid kinnitada – rakumembraani mikrograafidel olid selgelt näha lipiidide ja valgupeade kihid ning ka tühi ruum nende vahel.
1960. aastal töötas Ameerika bioloog J. Robertson välja teooria rakumembraanide kolmekihilise ehituse kohta, mida peeti pikka aega ainsaks tõeseks, kuid teaduse edasise arenguga hakkasid tekkima kahtlused selle eksimatus. Nii et näiteks rakkude seisukohalt oleks vajalike kasulike ainete transportimine läbi kogu “võileiva” keeruline ja töömahukas.
Ja alles 1972. aastal suutsid Ameerika bioloogid S. Singer ja G. Nicholson selgitada Robertsoni teooria ebakõlasid rakumembraani uue vedeliku-mosaiikmudeli abil. Eelkõige leidsid nad, et rakumembraan ei ole koostiselt homogeenne, pealegi on see asümmeetriline ja vedelikuga täidetud. Lisaks on rakud pidevas liikumises. Ja rakumembraani moodustavatel kurikuulsatel valkudel on erinev struktuur ja funktsioonid.

Rakumembraani omadused ja funktsioonid
Vaatame nüüd, milliseid funktsioone rakumembraan täidab:
Rakumembraani barjäärfunktsioon – membraan kui tõeline piirivalvur seisab valvel raku piiride üle, viivitab, ei lase läbi kahjulikke või lihtsalt sobimatuid molekule
Rakumembraani transpordifunktsioon - membraan ei ole ainult piirivalve raku väravates, vaid ka omamoodi tollipunkt, mida läbib pidevalt kasulike ainete vahetus teiste rakkude ja keskkonnaga.
Maatriksifunktsioon - see on rakumembraan, mis määrab üksteise suhtes asukoha, reguleerib nendevahelist interaktsiooni.
Mehaaniline funktsioon - vastutab ühe raku piiramise eest teisest ja paralleelselt rakkude õige ühendamise eest üksteisega, nende moodustumise eest homogeenseks koeks.
Rakumembraani kaitsefunktsioon on raku kaitsekilbi ehitamise aluseks. Looduses võib selle funktsiooni näideteks olla kõva puit, tihe nahk, kaitsekesta, kõik tänu membraani kaitsefunktsioonile.
Ensümaatiline funktsioon on veel üks oluline funktsioon, mida mõned rakuvalgud täidavad. Näiteks selle funktsiooni tõttu toimub seedeensüümide süntees sooleepiteelis.
Lisaks kõigele sellele toimub rakumembraani kaudu rakkude metabolism, mis võib toimuda kolme erineva reaktsioonina:
- Fagotsütoos on rakuvahetus, mille käigus membraani sisseehitatud fagotsüütrakud püüavad kinni ja seedivad erinevaid toitaineid.
- Pinotsütoos - on rakumembraani, sellega kokkupuutes olevate vedelikumolekulide kinnipüüdmise protsess. Selleks moodustuvad membraani pinnale spetsiaalsed kõõlused, mis justkui ümbritsevad vedelikutilka, moodustades mulli, mille membraan hiljem “alla neelab”.
- Eksotsütoos – on pöördprotsess, mille käigus rakk vabastab sekretoorse funktsionaalse vedeliku läbi membraani pinnale.

Rakumembraani struktuur
Rakumembraanis on kolm lipiidide klassi:
- fosfolipiidid (need on rasvade ja fosfori kombinatsioon),
- glükolipiidid (rasvade ja süsivesikute kombinatsioon),
- kolesterooli.
Fosfolipiidid ja glükolipiidid koosnevad omakorda hüdrofiilsest peast, millesse ulatuvad kaks pikka hüdrofoobset saba. Kolesterool seevastu hõivab nende sabade vahelise ruumi, takistades nende paindumist, kõik see muudab teatud rakkude membraani teatud juhtudel väga jäigaks. Lisaks kõigele sellele reguleerivad kolesterooli molekulid rakumembraani struktuuri.
Kuid olgu kuidas on, rakumembraani ehituse kõige olulisem osa on valk, õigemini erinevad valgud, mis täidavad erinevaid olulisi rolle. Vaatamata membraanis sisalduvate valkude mitmekesisusele, on midagi, mis neid ühendab – rõngakujulised lipiidid paiknevad kõigi membraanivalkude ümber. Rõngakujulised lipiidid on spetsiaalsed struktureeritud rasvad, mis toimivad valkude kaitsekestana, ilma milleta need lihtsalt ei töötaks.
Rakumembraani struktuur on kolmekihiline: rakumembraani aluseks on homogeenne vedel lipiidikiht. Valgud katavad seda mõlemalt poolt nagu mosaiik. Just valgud täidavad lisaks ülalkirjeldatud funktsioonidele ka omapäraste kanalite rolli, mille kaudu membraani läbivad ained, mis ei suuda membraani vedelat kihti tungida. Nende hulka kuuluvad näiteks kaaliumi- ja naatriumioonid, nende membraanist läbitungimiseks pakub loodus rakumembraanide spetsiaalseid ioonikanaleid. Teisisõnu tagavad valgud rakumembraanide läbilaskvuse.
Kui vaatame rakumembraani läbi mikroskoobi, näeme lipiidide kihti, mis on moodustunud väikestest sfäärilistest molekulidest, millel hõljuvad valgud nagu merel. Nüüd teate, millised ained on rakumembraani osa.
Rakumembraan, video
Ja lõpuks õpetlik video rakumembraani kohta.
9.5.1. Üks membraanide põhifunktsioone on osalemine ainete transpordis. Seda protsessi pakuvad kolm peamist mehhanismi: lihtne difusioon, hõlbustatud difusioon ja aktiivne transport (joonis 9.10). Pidage meeles nende mehhanismide kõige olulisemad omadused ja transporditavate ainete näited.
Joonis 9.10. Molekulide transportimise mehhanismid läbi membraani
lihtne difusioon- ainete ülekandmine läbi membraani ilma spetsiaalsete mehhanismide osaluseta. Transport toimub mööda kontsentratsioonigradienti ilma energiatarbimiseta. Väikesed biomolekulid - H2O, CO2, O2, uurea, hüdrofoobsed madala molekulmassiga ained transporditakse lihtsa difusiooni teel. Lihtsa difusiooni kiirus on võrdeline kontsentratsiooni gradiendiga.
Hõlbustatud difusioon- ainete ülekandmine läbi membraani valgukanalite või spetsiaalsete kandevalkude abil. See viiakse läbi piki kontsentratsioonigradienti ilma energiatarbimiseta. Transporditakse monosahhariide, aminohappeid, nukleotiide, glütserooli, mõningaid ioone. Iseloomulik on küllastuskineetika - ülekantava aine teatud (küllastava) kontsentratsiooni juures võtavad ülekandest osa kõik kandjamolekulid ja transpordikiirus jõuab piirväärtuseni.
aktiivne transport- nõuab ka spetsiaalsete kandevalkude osalemist, kuid ülekanne toimub kontsentratsioonigradienti vastu ja nõuab seetõttu energiat. Selle mehhanismi abil transporditakse läbi rakumembraani Na+, K+, Ca2+, Mg2+ ioonid, mitokondri membraani kaudu aga prootonid. Ainete aktiivset transporti iseloomustab küllastuskineetika.
9.5.2. Aktiivset ioonitransporti teostava transpordisüsteemi näide on Na+,K+ -adenosiintrifosfataas (Na+,K+ -ATPaas või Na+,K+ -pump). See valk asub plasmamembraani paksuses ja on võimeline katalüüsima ATP hüdrolüüsi reaktsiooni. 1 ATP molekuli hüdrolüüsil vabanevat energiat kasutatakse 3 Na + iooni kandmiseks rakust rakuvälisesse ruumi ja 2 K + iooni vastassuunas (joonis 9.11). Na +, K + -ATPaasi toime tulemusena tekib raku tsütosooli ja rakuvälise vedeliku kontsentratsioonide erinevus. Kuna ioonide transport on ebavõrdne, tekib elektripotentsiaalide erinevus. Seega tekib elektrokeemiline potentsiaal, mis on elektripotentsiaalide erinevuse energia Δφ ja ainete kontsentratsioonide erinevuse energia ΔС summa mõlemal pool membraani.

Joonis 9.11. Na+, K+ -pumba skeem.
9.5.3. Osakeste ja makromolekulaarsete ühendite ülekandmine läbi membraanide
Koos kandjate poolt läbiviidava orgaaniliste ainete ja ioonide transpordiga on rakus väga eriline mehhanism, mis on loodud makromolekulaarsete ühendite absorbeerimiseks ja rakust eemaldamiseks biomembraani kuju muutmise kaudu. Sellist mehhanismi nimetatakse vesikulaarne transport.

Joonis 9.12. Vesikulaarse transpordi tüübid: 1 - endotsütoos; 2 - eksotsütoos.
Makromolekulide ülekande käigus toimub membraaniga ümbritsetud vesiikulite (vesiikulite) järjestikune moodustumine ja sulandumine. Vastavalt transpordisuunale ja ülekantavate ainete olemusele eristatakse järgmisi vesikulaarse transpordi liike:
Endotsütoos(Joonis 9.12, 1) - ainete ülekandmine rakku. Sõltuvalt saadud vesiikulite suurusest on:
a) pinotsütoos - vedelate ja lahustunud makromolekulide (valgud, polüsahhariidid, nukleiinhapped) imendumine väikeste mullide (läbimõõduga 150 nm) abil;
b) fagotsütoos — suurte osakeste, nagu mikroorganismid või rakujäänused, imendumine. Sel juhul moodustuvad suured vesiikulid, mida nimetatakse fagosoomideks diameetriga üle 250 nm.
Pinotsütoos on iseloomulik enamikule eukarüootsetele rakkudele, samas kui suured osakesed imenduvad spetsialiseeritud rakkudesse - leukotsüüdid ja makrofaagid. Endotsütoosi esimeses etapis adsorbeeritakse ained või osakesed membraani pinnale, see protsess toimub ilma energiatarbimiseta. Järgmises etapis süveneb adsorbeeritud ainega membraan tsütoplasmasse; tekkivad plasmamembraani lokaalsed invaginatsioonid nööritakse rakupinnalt, moodustades vesiikulid, mis seejärel rändavad rakku. See protsess on ühendatud mikrokiudude süsteemiga ja on energiast sõltuv. Rakku sisenevad vesiikulid ja fagosoomid võivad lüsosoomidega ühineda. Lüsosoomides sisalduvad ensüümid lagundavad vesiikulites ja fagosoomides sisalduvad ained madala molekulmassiga saadusteks (aminohapped, monosahhariidid, nukleotiidid), mis transporditakse tsütosooli, kus rakk saab neid kasutada.
Eksotsütoos(Joonis 9.12, 2) - osakeste ja suurte ühendite ülekandmine rakust. See protsess, nagu endotsütoos, jätkub energia neeldumisega. Peamised eksotsütoosi tüübid on:
a) sekretsioon - kasutatavate või teisi keharakke mõjutavate veeslahustuvate ühendite eemaldamine rakust. Seda võivad läbi viia nii spetsialiseerimata rakud kui ka sisesekretsiooninäärmete rakud, seedetrakti limaskestad, mis on kohandatud nende poolt toodetavate ainete (hormoonid, neurotransmitterid, proensüümid) sekretsiooniks, olenevalt keha spetsiifilistest vajadustest. .
Sekreteeritud valgud sünteesitakse ribosoomidel, mis on seotud kareda endoplasmaatilise retikulumi membraanidega. Need valgud transporditakse seejärel Golgi aparaati, kus neid modifitseeritakse, kontsentreeritakse, sorteeritakse ja seejärel pakitakse vesiikulitesse, mis lõhustatakse tsütosooliks ja seejärel sulanduvad plasmamembraaniga nii, et vesiikulite sisu jääb rakust välja.
Erinevalt makromolekulidest transporditakse väikesed sekreteeritud osakesed, näiteks prootonid, rakust välja, kasutades hõlbustatud difusiooni ja aktiivseid transpordimehhanisme.
b) eritumist - ainete eemaldamine rakust, mida ei saa kasutada (näiteks erütropoeesi käigus retikulotsüütidest retikulaarse aine eemaldamine, mis on organellide agregeeritud jäänuk). Eritumise mehhanism seisneb ilmselt selles, et algul on eritunud osakesed tsütoplasmaatilises vesiikulis, mis seejärel sulandub plasmamembraaniga.
Kõik elusorganismid Maal koosnevad rakkudest ja iga rakku ümbritseb kaitsekesta – membraan. Kuid membraani funktsioonid ei piirdu ainult organellide kaitsmise ja ühe raku teisest eraldamisega. Rakumembraan on keeruline mehhanism, mis on otseselt seotud paljunemise, regeneratsiooni, toitumise, hingamise ja paljude teiste oluliste rakufunktsioonidega.
Mõistet "rakumembraan" on kasutatud umbes sada aastat. Sõna "membraan" tähendab ladina keelest "filmi". Aga rakumembraani puhul oleks õigem rääkida kahe teatud viisil omavahel ühendatud kile kombinatsioonist, pealegi on nende kilede erinevatel külgedel erinevad omadused.
Rakumembraan (tsütolemma, plasmalemma) on kolmekihiline lipoproteiini (rasvvalk) kest, mis eraldab iga raku naaberrakkudest ja keskkonnast ning teostab kontrollitud vahetust rakkude ja keskkonna vahel.
Selles määratluses ei ole määrav mitte see, et rakumembraan eraldaks ühe raku teisest, vaid see, et see tagab selle koostoime teiste rakkude ja keskkonnaga. Membraan on väga aktiivne, pidevalt töötav raku struktuur, millele on looduse poolt määratud palju funktsioone. Meie artiklist saate teada kõike rakumembraani koostise, struktuuri, omaduste ja funktsioonide kohta, samuti rakumembraanide talitlushäiretest tulenevate ohtude kohta inimeste tervisele.
Rakumembraanide uurimise ajalugu
1925. aastal suutsid kaks Saksa teadlast Gorter ja Grendel viia läbi kompleksse eksperimendi inimese punaste vereliblede ehk erütrotsüütidega. Osmootse šoki abil said teadlased nn "varjud" - punaste vereliblede tühjad kestad, seejärel panid need ühte hunnikusse ja mõõtsid pindala. Järgmine samm oli lipiidide hulga arvutamine rakumembraanis. Atsetooni abil eraldasid teadlased "varjudest" lipiidid ja tegid kindlaks, et neist piisab kahekordse pideva kihi jaoks.
Katse käigus tehti aga kaks jämedat viga:
Atsetooni kasutamine ei võimalda kõiki lipiide membraanidest eraldada;
"Varjude" pindala arvutati kuivkaalu järgi, mis on samuti vale.
Kuna esimene viga andis arvutustes miinuse, teine aga plussi, osutus üldtulemus üllatavalt täpseks ning Saksa teadlased tõid teadusmaailma olulisima avastuse - rakumembraani lipiidide kaksikkihi.
1935. aastal jõudis teine teadlaste paar Danielly ja Dawson pärast pikki katseid bilipiidkiledega järeldusele, et rakumembraanides leidub valke. Ei olnud muud võimalust seletada, miks neil kiledel on nii suur pindpinevus. Teadlased on avalikkuse ette toonud võileivalaadse rakumembraani skemaatilise mudeli, kus saiaviilude rolli täidavad homogeensed lipiid-valgukihid ning nende vahel on õli asemel tühjus.
1950. aastal leidis esimest elektronmikroskoobi kasutades Danielly-Dawsoni teooria osaliselt kinnitust – rakumembraani mikrograafidel olid selgelt näha kaks lipiidi- ja valgupeadest koosnevat kihti ning nende vahel oli läbipaistev ruum, mis oli täidetud ainult lipiidide sabadega. ja valgud.
1960. aastal töötas Ameerika mikrobioloog J. Robertson nendest andmetest juhindudes välja teooria rakumembraanide kolmekihilise struktuuri kohta, mida peeti pikka aega ainsaks tõeseks. Teaduse arenedes tekkis aga üha enam kahtlusi nende kihtide homogeensuses. Termodünaamika seisukohalt on selline struktuur äärmiselt ebasoodne - rakkudel oleks väga raske transportida aineid kogu “võileiva” kaudu sisse ja välja. Lisaks on tõestatud, et erinevate kudede rakumembraanid on erineva paksuse ja kinnitusviisiga, mis on tingitud elundite erinevatest funktsioonidest.
1972. aastal leidsid mikrobioloogid S.D. Laulja ja G.L. Nicholson suutis kõik Robertsoni teooria ebakõlad selgitada rakumembraani uue vedeliku-mosaiikmudeli abil. Teadlased on leidnud, et membraan on heterogeenne, asümmeetriline, vedelikuga täidetud ja selle rakud on pidevas liikumises. Ja selle moodustavad valgud on erineva struktuuri ja eesmärgiga, lisaks paiknevad nad membraani bilipiidkihi suhtes erinevalt.
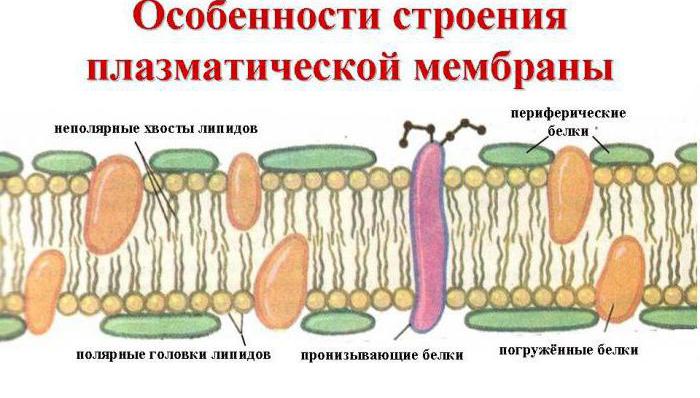
Rakumembraanid sisaldavad kolme tüüpi valke:
Perifeerne - kinnitatud kile pinnale;
poolintegraalne- tungida osaliselt läbi bilipiidkihi;
Integraalne - tungib täielikult läbi membraani.
Perifeersed valgud on elektrostaatilise interaktsiooni kaudu seotud membraani lipiidide peadega ja nad ei moodusta kunagi pidevat kihti, nagu varem arvati. Ja poolintegraalsed ja integraalsed valgud transpordivad rakku hapnikku ja toitaineid, samuti eemaldavad lagunemise. selle tooteid ja palju muud mitmete oluliste funktsioonide jaoks, millest saate hiljem teada.

Rakumembraan täidab järgmisi funktsioone:
Barjäär – membraani läbilaskvus erinevat tüüpi molekulide puhul ei ole ühesugune.Rakumembraanist möödasõiduks peab molekul olema kindla suurusega, keemilised omadused ja elektrilaeng. Kahjulikud või sobimatud molekulid ei saa rakumembraani barjäärifunktsiooni tõttu lihtsalt rakku siseneda. Näiteks kaitseb membraan peroksiidreaktsiooni abil tsütoplasmat talle ohtlike peroksiidide eest;
Transport – membraani läbib passiivne, aktiivne, reguleeritud ja selektiivne vahetus. Passiivne ainevahetus sobib väga väikestest molekulidest koosnevatele rasvlahustuvatele ainetele ja gaasidele. Sellised ained tungivad difusiooni teel vabalt rakku ja sealt välja ilma energiakuluta. Vajadusel aktiveeritakse rakumembraani aktiivne transpordifunktsioon, kuid raskesti transporditavaid aineid on vaja rakku sisse või sealt välja viia. Näiteks need, kellel on suur molekulaarne suurus või mis ei suuda hüdrofoobsuse tõttu bilipiidkihti ületada. Seejärel hakkavad tööle valgupumbad, sealhulgas ATPaas, mis vastutab kaaliumiioonide imendumise eest rakku ja naatriumioonide väljutamise eest. Reguleeritud transport on oluline sekretsiooni ja fermentatsiooni funktsioonide jaoks, näiteks kui rakud toodavad ja eritavad hormoone või maomahla. Kõik need ained lahkuvad rakkudest spetsiaalsete kanalite kaudu ja etteantud mahus. Ja selektiivne transpordifunktsioon on seotud väga integreeritud valkudega, mis tungivad läbi membraani ja toimivad kanalina rangelt määratletud tüüpi molekulide sisenemiseks ja väljumiseks;
Maatriks – rakumembraan määrab ja fikseerib organellide paiknemise üksteise suhtes (tuum, mitokondrid, kloroplastid) ning reguleerib nendevahelist vastasmõju;
Mehaaniline - tagab ühe raku piiramise teisest ja samal ajal rakkude õige ühendamise homogeenseks koeks ja elundite vastupidavuse deformatsioonile;
Kaitsev - nii taimedes kui loomades on rakumembraan kaitseraami ehitamise aluseks. Näiteks kõva puit, tihe koor, torkivad okkad. Loomamaailmas on ka palju näiteid rakumembraanide kaitsefunktsioonist – kilpkonna kest, kitiinkarp, kabjad ja sarved;
Energia - fotosünteesi ja rakuhingamise protsessid oleksid võimatud ilma rakumembraani valkude osaluseta, sest rakud vahetavad energiat just valgukanalite abil;
Retseptor – rakumembraani põimitud valkudel võib olla veel üks oluline funktsioon. Need toimivad retseptoritena, mille kaudu rakk saab signaali hormoonidelt ja neurotransmitteritelt. Ja see on omakorda vajalik närviimpulsside juhtimiseks ja hormonaalsete protsesside normaalseks kulgemiseks;
Ensümaatiline - veel üks oluline funktsioon, mis on omane mõnedele rakumembraanide valkudele. Näiteks sooleepiteelis sünteesitakse selliste valkude abil seedeensüüme;
Biopotentsiaal- kaaliumiioonide kontsentratsioon rakus on palju suurem kui väljaspool ja naatriumiioonide kontsentratsioon, vastupidi, on suurem väljaspool kui sees. See seletab potentsiaalset erinevust: laeng on negatiivne rakus sees, positiivne väljaspool, mis aitab kaasa ainete liikumisele rakku ja sealt välja mis tahes kolme tüüpi ainevahetuses – fagotsütoos, pinotsütoos ja eksotsütoos;
Märgistus - rakumembraanide pinnal on nn "märgised" - glükoproteiinidest koosnevad antigeenid (valgud, mille külge on kinnitatud hargnenud oligosahhariidsed kõrvalahelad). Kuna külgahelatel võib olla väga erinevaid konfiguratsioone, saab iga rakutüüp oma ainulaadse märgise, mis võimaldab teistel keharakkudel neid "nägemise järgi" ära tunda ja neile õigesti reageerida. Seetõttu tunnevad näiteks inimese immuunrakud, makrofaagid, kehasse sattunud välismaalase (infektsioon, viirus) kergesti ära ja püüavad seda hävitada. Sama juhtub haigete, muteerunud ja vanade rakkudega – nende rakumembraanil muutub silt ja keha vabaneb neist.
Rakuvahetus toimub läbi membraanide ja seda saab läbi viia kolme peamise reaktsioonitüübi kaudu:
Fagotsütoos on rakuline protsess, mille käigus membraani sisseehitatud fagotsüütrakud püüavad kinni ja seedivad toitainete tahkeid osakesi. Inimkehas viivad fagotsütoosi läbi kahte tüüpi rakkude membraanid: granulotsüüdid (granuleeritud leukotsüüdid) ja makrofaagid (immuunsuse tapjarakud);
Pinotsütoos on vedelate molekulide hõivamine, mis puutuvad sellega kokku rakumembraani pinna kaudu. Pinotsütoosi tüübi järgi toitumiseks kasvatab rakk oma membraanile õhukesed kohevad väljakasvud antennide kujul, mis ümbritsevad justkui vedelikutilka, ja tekib mull. Esiteks ulatub see vesiikul membraani pinnast kõrgemale ja seejärel "alla neelatakse" - see peidab end raku sees ja selle seinad ühinevad rakumembraani sisepinnaga. Pinotsütoos esineb peaaegu kõigis elusrakkudes;
Eksotsütoos on pöördprotsess, mille käigus tekivad raku sees vesiikulid sekretoorse funktsionaalse vedelikuga (ensüüm, hormoon), mis tuleb kuidagi rakust keskkonda eemaldada. Selleks sulandub mull esmalt rakumembraani sisepinnaga, seejärel pundub väljapoole, lõhkeb, ajab sisu välja ja sulandub uuesti membraani pinnaga, seekord väljastpoolt. Eksotsütoos toimub näiteks sooleepiteeli ja neerupealiste koore rakkudes.

Rakumembraanid sisaldavad kolme lipiidide klassi:
fosfolipiidid;
glükolipiidid;
Kolesterool.
Fosfolipiidid (rasvade ja fosfori kombinatsioon) ja glükolipiidid (rasvade ja süsivesikute kombinatsioon) koosnevad omakorda hüdrofiilsest peast, millest ulatub välja kaks pikka hüdrofoobset saba. Kuid kolesterool hõivab mõnikord nende kahe saba vahelise ruumi ega lase neil painduda, mis muudab mõne raku membraanid jäigaks. Lisaks muudavad kolesterooli molekulid sujuvamaks rakumembraanide struktuuri ja takistavad polaarsete molekulide üleminekut ühest rakust teise.
Kuid kõige olulisem komponent, nagu võib näha eelmisest rakumembraanide funktsioone käsitlevast jaotisest, on valgud. Nende koostis, otstarve ja asukoht on väga mitmekesised, kuid on midagi ühist, mis neid kõiki ühendab: rõngakujulised lipiidid paiknevad alati rakumembraanide valkude ümber. Need on spetsiaalsed rasvad, mis on selgelt struktureeritud, stabiilsed, nende koostises on rohkem küllastunud rasvhappeid ja mis vabanevad membraanidest koos "sponsoreeritud" valkudega. See on omamoodi isiklik kaitsekesta valkude jaoks, ilma milleta need lihtsalt ei töötaks.
Rakumembraani struktuur on kolmekihiline. Keskel on suhteliselt homogeenne vedel bilipiidkiht ja valgud katavad seda mõlemalt poolt omamoodi mosaiigiga, tungides osaliselt paksusesse. See tähendab, et oleks vale arvata, et rakumembraanide välimised valgukihid on pidevad. Valgud on lisaks nende keerukatele funktsioonidele vajalikud membraanis selleks, et rakkudesse siseneda ja sealt välja transportida need ained, mis ei suuda rasvakihti tungida. Näiteks kaaliumi- ja naatriumioonid. Nende jaoks on ette nähtud spetsiaalsed valgustruktuurid - ioonkanalid, mida käsitleme üksikasjalikumalt allpool.
Kui vaadata rakumembraani läbi mikroskoobi, on näha kõige väiksematest sfäärilistest molekulidest moodustunud lipiidide kihti, mida mööda hõljuvad sarnaselt merega suured erineva kujuga valgurakud. Täpselt samad membraanid jagavad iga raku siseruumi osadeks, milles tuum, kloroplastid ja mitokondrid mugavalt paiknevad. Kui raku sees ei oleks eraldi “ruume”, kleepuksid organellid kokku ega suudaks oma funktsioone õigesti täita.
Rakk on membraanidega struktureeritud ja piiritletud organellide kogum, mis osaleb energia-, ainevahetus-, informatsiooni- ja paljunemisprotsesside kompleksis, mis tagavad organismi elutegevuse.
Nagu sellest määratlusest näha, on membraan iga raku kõige olulisem funktsionaalne komponent. Selle tähtsus on sama suur kui tuuma, mitokondrite ja teiste rakuorganellide oma. Ja membraani ainulaadsed omadused tulenevad selle struktuurist: see koosneb kahest erilisel viisil kokku kleepunud kilest. Fosfolipiidide molekulid membraanis paiknevad hüdrofiilsete peadega väljapoole ja hüdrofoobsete sabadega sissepoole. Seetõttu on kile üks pool veest märjaks, teine aga mitte. Niisiis on need kiled üksteisega ühendatud mittemärguvate külgedega sissepoole, moodustades bilipiidkihi, mida ümbritsevad valgumolekulid. See on rakumembraani "sandwich" struktuur.
Rakumembraanide ioonikanalid
Vaatleme üksikasjalikumalt ioonkanalite tööpõhimõtet. Milleks neid vaja on? Fakt on see, et ainult rasvlahustuvad ained võivad vabalt läbi lipiidmembraani tungida - need on gaasid, alkoholid ja rasvad ise. Nii näiteks toimub punastes verelibledes pidev hapniku ja süsihappegaasi vahetus ning selleks ei pea meie keha kasutama mingeid täiendavaid nippe. Aga mis siis, kui on vaja transportida vesilahuseid, näiteks naatriumi- ja kaaliumisoolasid, läbi rakumembraani?
Sellistele ainetele oleks bilipiidkihis võimatu teed sillutada, kuna augud tõmbuksid kohe kokku ja kleepuksid kokku tagasi, selline on iga rasvkoe struktuur. Kuid loodus, nagu alati, leidis olukorrast väljapääsu ja lõi spetsiaalsed valgu transpordistruktuurid.
Juhtivaid valke on kahte tüüpi:
Transporterid on poolintegreeritud valgupumbad;
Kanaloformerid on lahutamatud valgud.
Esimest tüüpi valgud on osaliselt sukeldatud rakumembraani bilipiidkihti ja vaatavad oma peaga välja ning soovitud aine juuresolekul hakkavad nad käituma nagu pump: meelitavad molekuli ja imevad selle endasse. kamber. Ja teist tüüpi, integraalsed valgud on pikliku kujuga ja asuvad rakumembraani bilipiidkihiga risti, tungides selle läbi ja läbi. Nende kaudu, nagu ka tunnelite kaudu, liiguvad rakku sisse ja sealt välja ained, mis ei suuda rasvast läbi minna. Ioonikanalite kaudu tungivad kaaliumiioonid rakku ja kogunevad sinna, naatriumioonid aga vastupidi. Elektrilistes potentsiaalides on erinevus, mis on nii vajalik meie keha kõigi rakkude nõuetekohaseks toimimiseks.
Olulisemad järeldused rakumembraanide ehituse ja funktsioonide kohta

Teooria tundub alati huvitav ja paljutõotav, kui seda saab praktikas kasulikult rakendada. Inimkeha rakumembraanide struktuuri ja funktsioonide avastamine võimaldas teadlastel teha tõelise läbimurde teaduses üldiselt ja eriti meditsiinis. Pole juhus, et oleme ioonkanalitel nii üksikasjalikult peatunud, sest just siin peitub vastus meie aja ühele kõige olulisemale küsimusele: miks haigestuvad inimesed üha sagedamini onkoloogiasse?
Vähk nõuab igal aastal maailmas umbes 17 miljonit inimelu ja on kõigi surmade põhjuste hulgas neljas. WHO andmetel kasvab vähki haigestumine pidevalt ning 2020. aasta lõpuks võib see ulatuda 25 miljonini aastas.
Mis seletab tõelist vähiepideemiat ja kuidas on sellega pistmist rakumembraanide funktsioon? Te ütlete: põhjuseks on halvad keskkonnatingimused, alatoitumus, halvad harjumused ja raske pärilikkus. Ja muidugi on teil õigus, aga kui probleemist täpsemalt rääkida, siis põhjuseks on inimkeha hapestumine. Eespool loetletud negatiivsed tegurid põhjustavad rakumembraanide häireid, pärsivad hingamist ja toitumist.
Seal, kus peaks olema pluss, tekib miinus ja rakk ei saa normaalselt toimida. Kuid vähirakud ei vaja ei hapnikku ega leeliselist keskkonda - nad on võimelised kasutama anaeroobset tüüpi toitumist. Seetõttu muteeruvad terved rakud hapnikuvaeguse ja skaalavälise pH-taseme tingimustes, soovides keskkonnaga kohaneda ja muutuvad vähirakkudeks. Nii haigestub inimene vähki. Selle vältimiseks peate lihtsalt iga päev jooma piisavalt puhast vett ja loobuma kantserogeenidest toidus. Kuid reeglina on inimesed kahjulikest toodetest ja kvaliteetse vee vajadusest teadlikud ega tee midagi - nad loodavad, et hädad lähevad neist mööda.
Teades erinevate rakkude rakumembraanide struktuuri ja funktsioonide omadusi, saavad arstid seda teavet kasutada, et pakkuda kehale sihipärast ja sihipärast ravitoimet. Paljud kaasaegsed ravimid, sattudes meie kehasse, otsivad õiget "sihtmärki", milleks võivad olla rakumembraanide ioonkanalid, ensüümid, retseptorid ja biomarkerid. See ravimeetod võimaldab saavutada paremaid tulemusi minimaalsete kõrvalmõjudega.
Viimase põlvkonna antibiootikumid ei tapa verre sattudes kõiki rakke järjest, vaid otsivad täpselt üles patogeeni rakud, keskendudes selle rakumembraanides olevatele markeritele. Uusimad migreenivastased ravimid, triptaanid, ahendavad ainult põletikulisi aju veresooni, avaldamata peaaegu mingit mõju südamele ja perifeersele vereringesüsteemile. Ja nad tunnevad vajalikud veresooned ära täpselt oma rakumembraanide valkude järgi. Selliseid näiteid on palju, seega võime kindlalt väita, et teadmised rakumembraanide ehitusest ja funktsioonidest on tänapäevase arstiteaduse arengu aluseks ning päästavad igal aastal miljoneid elusid.
Haridus: Moskva meditsiiniinstituut. I. M. Sechenov, eriala - "Meditsiin" 1991. aastal, 1993. aastal "Kutsehaigused", 1996. aastal "Teraapia".
Organismide, aga ka taimede, loomade ja inimeste ehituse uurimine on bioloogia haru, mida nimetatakse tsütoloogiaks. Teadlased on leidnud, et selle sees oleva raku sisu on üsna keeruline. Seda ümbritseb nn pinnaaparaat, mis hõlmab raku välismembraani, membraaniüleseid struktuure: glükokalüks ja mikrofilamendid, peliikulid ja mikrotuubulid, mis moodustavad selle submembraanse kompleksi.
Selles artiklis uurime välise rakumembraani struktuuri ja funktsioone, mis on osa erinevat tüüpi rakkude pinnaaparaadist.
Millised on raku välismembraani funktsioonid?
Nagu varem kirjeldatud, on välismembraan iga raku pinnaaparaadi osa, mis eraldab edukalt selle sisemise sisu ja kaitseb rakuorganelle ebasoodsate keskkonnatingimuste eest. Teiseks funktsiooniks on tagada ainete vahetus rakusisu ja koevedeliku vahel, seetõttu transpordib raku välimine membraan tsütoplasmasse sisenevaid molekule ja ioone, samuti aitab rakust välja viia toksiine ja liigseid mürgiseid aineid.

Rakumembraani struktuur
Erinevat tüüpi rakkude membraanid ehk plasmalemmad on üksteisest väga erinevad. Peamiselt keemiline struktuur, samuti lipiidide, glükoproteiinide, valkude suhteline sisaldus neis ja vastavalt ka neis olevate retseptorite olemus. Väline, mille määrab peamiselt glükoproteiinide individuaalne koostis, osaleb keskkonna stiimulite äratundmises ja raku enda reaktsioonides nende tegevusele. Teatud tüüpi viirused võivad interakteeruda rakumembraanide valkude ja glükolipiididega, mille tulemusena nad tungivad rakku. Herpes- ja gripiviirused võivad oma kaitsekesta ehitada.
Ja viirused ja bakterid, nn bakteriofaagid, kinnituvad rakumembraanile ja lahustavad selle kokkupuutepunktis spetsiaalse ensüümi abil. Seejärel liigub moodustunud auku viiruse DNA molekul.
Eukarüootide plasmamembraani struktuuri tunnused
Tuletage meelde, et raku välimine membraan täidab transpordi funktsiooni, see tähendab ainete ülekandmist sellesse ja sealt välja väliskeskkonda. Sellise protsessi läbiviimiseks on vaja spetsiaalset struktuuri. Tõepoolest, plasmalemma on kõigi jaoks pidev ja universaalne pinnaaparaadi süsteem. See on õhuke (2-10 Nm), kuid üsna tihe mitmekihiline kile, mis katab kogu raku. Selle struktuuri uurisid 1972. aastal sellised teadlased nagu D. Singer ja G. Nicholson, nad lõid ka rakumembraani vedeliku-mosaiikmudeli.
Peamised seda moodustavad keemilised ühendid on valkude ja teatud fosfolipiidide järjestatud molekulid, mis paiknevad vedelas lipiidkeskkonnas ja meenutavad mosaiiki. Seega koosneb rakumembraan kahest lipiidide kihist, mille mittepolaarsed hüdrofoobsed "sabad" asuvad membraani sees ning polaarsed hüdrofiilsed pead on suunatud raku tsütoplasma ja interstitsiaalse vedeliku poole.
Lipiidikihti tungivad läbi suured valgumolekulid, mis moodustavad hüdrofiilseid poore. Nende kaudu transporditakse glükoosi ja mineraalsoolade vesilahuseid. Mõned valgumolekulid paiknevad nii plasmalemma välis- kui ka sisepinnal. Seega on kõigi tuumadega organismide rakkude välisel rakumembraanil süsivesikute molekulid, mis on seotud kovalentsete sidemetega glükolipiidide ja glükoproteiinidega. Süsivesikute sisaldus rakumembraanides on vahemikus 2 kuni 10%.

Prokarüootsete organismide plasmalemma struktuur
Välimine rakumembraan täidab prokarüootidel sarnaseid funktsioone tuumaorganismide rakkude plasmamembraanidega, nimelt: väliskeskkonnast tuleva teabe tajumine ja edastamine, ioonide ja lahuste transport rakku ja sealt välja ning raku kaitsmine. tsütoplasma välistest reaktiividest. See võib moodustada mesosoome - struktuure, mis tekivad siis, kui plasmalemma tungib rakku. Need võivad sisaldada ensüüme, mis osalevad prokarüootide metaboolsetes reaktsioonides, näiteks DNA replikatsioonis, valkude sünteesis.
Mesosoomid sisaldavad ka redoksensüüme, fotosünteetikumid aga bakterioklorofülli (bakterites) ja fükobiliini (tsüanobakterites).
Välismembraanide roll rakkudevahelistes kontaktides
Jätkates vastamist küsimusele, milliseid funktsioone täidab välimine rakumembraan, peatume pikemalt selle rollil taimerakkudes Taimerakkudes tekivad rakuvälise membraani seintesse poorid, mis lähevad tselluloosikihti. Nende kaudu on võimalik raku tsütoplasma väljumine väljapoole, selliseid õhukesi kanaleid nimetatakse plasmodesmaatideks.

Tänu neile on ühendus naabertaimerakkude vahel väga tugev. Inimese ja looma rakkudes nimetatakse külgnevate rakumembraanide kokkupuutekohti desmosoomideks. Need on iseloomulikud endoteeli- ja epiteelirakkudele ning neid leidub ka kardiomüotsüütides.
Plasmalemma abimoodustised
Et mõista, kuidas taimerakud loomadest erinevad, aitab see uurida nende plasmamembraanide struktuurseid iseärasusi, mis sõltuvad sellest, milliseid funktsioone raku välimine membraan täidab. Loomarakkudes on selle kohal glükokalüksi kiht. Selle moodustavad polüsahhariidimolekulid, mis on seotud raku välismembraani valkude ja lipiididega. Tänu glükokalüksile toimub rakkude vahel adhesioon (kleepumine), mis viib kudede moodustumiseni, seetõttu osaleb see plasmalemma signaalimisfunktsioonis - keskkonna stiimulite äratundmises.
Kuidas toimub teatud ainete passiivne transport läbi rakumembraanide
Nagu varem mainitud, osaleb raku välimine membraan ainete transportimise protsessis raku ja väliskeskkonna vahel. Plasmalemma kaudu on kahte tüüpi transporti: passiivne (difusioon) ja aktiivne transport. Esimene hõlmab difusiooni, hõlbustatud difusiooni ja osmoosi. Ainete liikumine piki kontsentratsioonigradienti sõltub eelkõige rakumembraani läbivate molekulide massist ja suurusest. Näiteks väikesed mittepolaarsed molekulid lahustuvad kergesti plasmalemma keskmises lipiidikihis, liiguvad sellest läbi ja satuvad tsütoplasmasse.

Suured orgaaniliste ainete molekulid tungivad spetsiaalsete kandevalkude abil tsütoplasmasse. Need on liigispetsiifilised ja koos osakese või iooniga kannavad need passiivselt läbi membraani mööda kontsentratsioonigradienti ilma energiat kulutamata (passiivne transport). Selle protsessi aluseks on plasmalemma selline omadus nagu selektiivne läbilaskvus. Protsessis ei kasutata ATP molekulide energiat ning rakk säästab seda muudeks metaboolseteks reaktsioonideks.
Keemiliste ühendite aktiivne transport läbi plasmalemma
Kuna raku välimine membraan tagab molekulide ja ioonide ülekande väliskeskkonnast rakku ja tagasi, on võimalik eemaldada dissimilatsiooniproduktid, mis on toksiinid, väljapoole ehk rakkudevahelisse vedelikku. toimub kontsentratsioonigradiendi vastu ja nõuab energia kasutamist ATP molekulide kujul. See hõlmab ka kandevalke, mida nimetatakse ATPaasideks, mis on samuti ensüümid.

Sellise transpordi näiteks on naatrium-kaaliumpump (naatriumioonid liiguvad tsütoplasmast väliskeskkonda ja kaaliumiioonid pumbatakse tsütoplasmasse). Soole ja neerude epiteelirakud on selleks võimelised. Selle ülekandemeetodi variandid on pinotsütoosi ja fagotsütoosi protsessid. Seega, olles uurinud, milliseid funktsioone raku välimine membraan täidab, saab kindlaks teha, et heterotroofsed protistid, aga ka kõrgemate loomorganismide rakud, näiteks leukotsüüdid, on võimelised pino- ja fagotsütoosiks.
Bioelektrilised protsessid rakumembraanides
On kindlaks tehtud, et plasmalemma välispinna (see on positiivselt laetud) ja tsütoplasma parietaalkihi vahel, mis on negatiivselt laetud, on potentsiaalide erinevus. Seda nimetati puhkepotentsiaaliks ja see on omane kõigile elusrakkudele. Ja närvikoel pole mitte ainult puhkepotentsiaali, vaid see on võimeline juhtima ka nõrku biovoolusid, mida nimetatakse ergastusprotsessiks. Närvirakkude-neuronite välismembraanid, mis saavad retseptoritelt ärritust, hakkavad muutma laenguid: naatriumioonid sisenevad massiliselt rakku ja plasmalemma pind muutub elektronegatiivseks. Ja tsütoplasma parietaalne kiht saab katioonide liigse tõttu positiivse laengu. See seletab, miks neuroni välimine rakumembraan laetakse uuesti, mis põhjustab ergastusprotsessi aluseks olevate närviimpulsside juhtivuse.
Seotud väljaanded
-
 Unenägude tähendus Natalia Stepanova suure unenägude raamatu järgi
Unenägude tähendus Natalia Stepanova suure unenägude raamatu järgi
Teen ettepaneku mitte seletada traditsioonilises mõttes unenägusid mitte analüüsida, vaid neid tõlgendada, nagu inimesed teevad ja on teinud ...
-
 Suurimad peamised naftajuhtmed
Suurimad peamised naftajuhtmed
Nafta ja naftasaaduste meretranspordiks mõeldud maailma tankerlaevastik erineb teistest kaubalaevanduse sektoritest ...